Carbohydrate Research (1998) 306, 335-339.
Department of Chemistry & Biochemistry,
University of Maryland Baltimore County, Baltimore, MD 21228.
Research supported by NSF Grant MCB 91-05586 Running title: C-C Coupling in Polysaccharides * Corresponding author - ph. (410) 455-2506, fax (410) 455-2608, e-mail bush@umbc.edu
Keywords: polysaccharide conformtion, carbon-carbon coupling, NMR spectroscopy, oligosaccharide
1. Introduction
Factors influencing the conformation and the dynamics of complex carbohydrate epitopes are not completely understood. Although some workers have proposed that the conformations are relatively rigid and are imposed by stereochemical interactions [1,2,3], others have emphasized the flexible nature of the oligosaccharide structures [4,5]. Most conformational models of complex oligosaccharides are derived from NOE data in NMR spectroscopy and oligosaccharide models based on a few NOE distance constraints combined with computer molecular modeling have been satisfactory for molecules which are reasonably rigid [6,7,8]. This treatment has been less successful for more flexible molecules [9].
Scalar coupling data can be a valuable complement to the NOE in oligosaccharides since the J values between atoms across the glycosidic linkage report directly on the glycosidic dihedral angles. J coupling has additional advantages in averaging over multiple conformations of flexible oligosaccharides. For NOE data, averaging over multiple conformations is complicated both by the 1/r6 dependance on distance and by the complex and poorly understood time dependance in which the overall molecular tumbling may interact with kinetics of the conformational exchange involved in the internal motion. Averaging of scalar coupling values over multiple conformations is a simple linear average over individual conformers.
The potential value of 13C13 coupling data between atoms on opposite sides of the glycosidic linkage has long been recognized, [10,11] and trigonometric correlation curves relating 3JCH to the glycosidic dihedral angles, F and Y have been proposed [12,13]. 3JCH data have been measured for complex oligosaccharides studied in natural abundance 13C13 using indirect detection [14,15,16,17]. With complex polysaccharides enriched in 13C13, a large number of relevant coupling constants can be readily measured for high molecualr weight polysaccharides with good accuracy, even for small values of 3JCH [18,19,20].
As a complement to 3JCH data, nJCC could provide information useful in resolving the ambiguity resulting from the multi-valued coupling constant correlation curves as well as more data to determine statistical weights of multiple conformations in flexible models. nJCC values have been reported in monosaccharides and simple disaccharides [21,22,23] but no experimental method has been described suitable for high molecular weight polysaccharides or complex oligosaccharides. In the present communication, we describe the application of a method reported by Bax, Max and Zax [24] for 1H1 detected measurement of 3JCC of methyl resonances in proteins to complex carbohydrates including high molecular weight complex polysaccharides.
II. Materials and Methods
Uniformly 13C13 enriched glucose was purchased from Isotec. The cell wall lectin receptor polysaccharide from Streptococcus mitis J22 was extracted from bacteria grown in media uniformly highly enriched in 13C13 and the product polysaccharide was labeled to an extent of about 96% 13C13. [18]. This polysaccharide (Fig. 1) having seven sugar residues in the repeating subunit [25] was dissolved in D2O2o at a concentration of 8 mg/ml corresponding to an approximate concentration of 8 mM. NMR spectra were recorded on a GE Omega 600 spectrometer using a Bruker triple resonance probe at 24° C . Data were recorded with the pulse sequence of Bax et al. [24]. Short 13C13 90°pulse lengths (15 msec in these experiments) are important for this experiment in order to excite coupled carbon atoms with uniform power and phase. The delay, T, was set to correspond to 44 or 46 Hz to minimize the influence of 1JCC and the delay, t, for transfer of magnetization by 1JCH was set to correspond to 168 Hz. Coupling constants between Ca and Cb were extracted, using eq. 1, from the ratio of the cross peak between the proton Ha (attached to Ca) and Cb and the direct peak between Ha and Ca.[24]
| (1) |
III. Results and Discussion
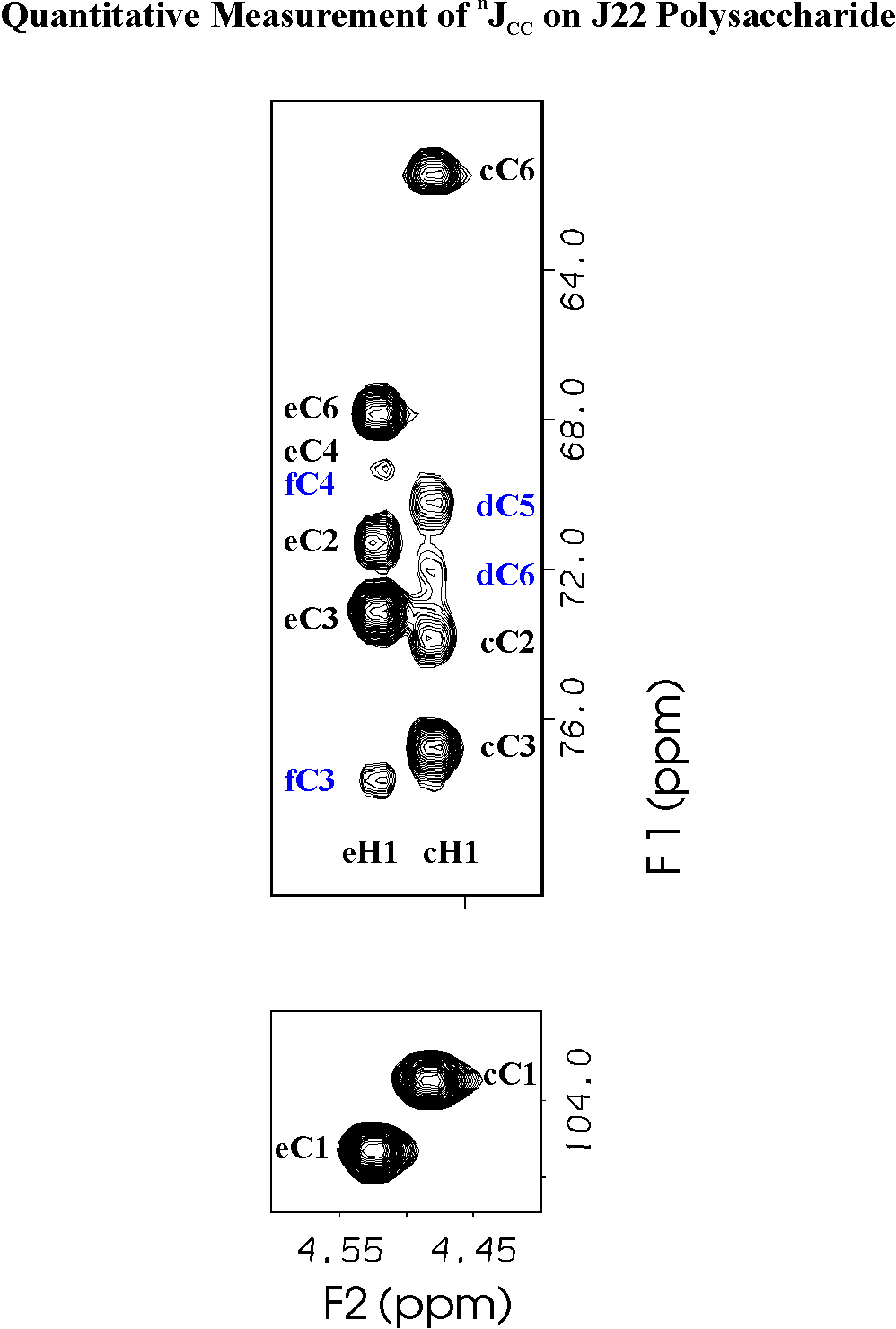
The pulse sequence used gives a 2-d C-H correlation spectrum in which the magnitude of a cross peak to a 13C13 resonance is related by eq. 1 to the nJCC between that carbon atom and the carbon atom attached to the proton in the column. The large peak between the proton and its directly bonded 13C13 is used as a reference peak in the calculation and the effect of the one-bond 13C13-13C13 coupling to the adjacent carbon atom is minimized by choice of the delay period, T, in the pulse sequence of Bax et al [24]. Fig. 2 shows 1H1 signals from two anomeric proton signals with cross peaks to carbon atoms of the same residue and to the carbon atoms of the succeeding residue of the polysaccharide sequence.
As a control on the precision and accuracy of our measurements of the nJCC , we have recorded data for uniformly enriched glucose for comparison with values reported previously by Serianni and coworkers [21,22], who measured splittings in direct 13C13 detected spectra of monosaccharides enriched with 13C13 at single sites. For glucose, (Table 1.) our measured values agree within the stated experimental error (±0.3 Hz).
For the cell wall polysaccharide of S. mitis J22, the residues of which are identified by letters in structure of Fig. 1, measurements of intraring 3JCC are expected to be very similar to values reported for monosaccharides since they depend primarily on puckering of the sugar ring. Data in Table 2 show that the measured values for residue c agree with values reported for b-glc and for residue e agree with published data for b-gal. nJCC measured for residues f and a (both a-galNAc ) agree well with data reported for a-gal suggesting only a small effect of the NAc substituent at C2. Measured values for residues b and g (b-and a-rhamnose respectively) agree with reported values for b-and a-mannose with the exception of our measured 2JCC for b C1-C3 (5.0 Hz) which differs substantially from the corresponding value (4.0 Hz) for b-man. It is possible that the failure of some of the intraring nJCC measured for the polysaccharide to agree with those for isolated monosaccharides could result from effects of substituents in the polymer.
Several types of cross peaks relevant to the conformation of polysaccharides are seen in the spectrum. In Fig. 2 on the column of the anomeric 1H1 signal, cross peaks are seen representing 3JCC between C1 and Cx±1 which are related to the glycosidic dihedral angle, Y. Most of these cross peaks can be detected and are reported in Table 2. The technical problems of measuring the relevant coupling constants differ somewhat from those of methyl groups of protein side chains reported by Bax et al [24] as a result of differences in the chemical shifts. While the most accurate quantitation of this coupling constant can be determined from the column corresponding to the anomeric proton signal, any small residual antiphase magnetization in the spectrum can make it difficult to phase the spectrum correctly if the coupling constant is smaller than about 1.5 Hz. The same coupling constant can also be measured on the columns of Hx±1 (data not shown). Unfortunately, as a result of the poor chemical shift dispersion of carbohydrates, this isolated C1-Hx cross peak cannot be easily interpreted if the signal of Hx overlaps with that of another proton whose attached carbon could be coupled to C1. But the cross peak from Hx±1 to C1 is in an uncrowded region and can be accurately phased to ascertain whether the nJCC value is greater than 1 Hz or is truly zero. The nJCC values greater than 1 Hz in Table 1 have a precision of ± 0.3 Hz while values lower than 1 Hz are less precise because the peak becomes comparable to noise due to the tan2 relationship in the trigonometric formula (eq. 1) which relates cross peak intensity to nJCC.
While 3JC2-CX, related to the glycosidic dihedral angle F are potentially valuable, their measurement is complicated by overlap of the signals of H2, Hx, C2 and Cx which are often in crowded regions of the spectrum but four of the values are reported in Table 1. It has been recently proposed that the two-bond coupling, 2JC1-CX, is related to this same dihedral angle [26,27]. The magnitudes, but not the signs, of these coupling constants are reported in Table 1.
The values of 3JCC are related to the glycosidic dihedral angles by a trigonometric relation of the form of Eq. 2.
| (2) |
In Table 2, we include values of 3JCC calculated from the flexible model for the J22 polysaccharide proposed earlier [19]. There is some disagreement between experiment and values calculated from the model which includes statistical weights of three conformations. Several of the calculated values lie well outside experimental error and the discrepancy is of a fairly fundamental nature. For the glycosidic dihedral angles Yab and Ybc between the a-galNAc and b-rha residue and between the b-rha and b-glc residue, the reported values of 3JCH are small (1.5 and 2.1 Hz respectively), [19] implying that C1 is nearly gauche to the aglycone proton. Thus, for any single conformation, C1 must be nearly trans to either Cx+1 or to Cx-1 but in fact the data of Table 2 show both 3JCC values to be much less than 4 Hz. Either the coupling constant correlation predicts values of 3JCH and 3JCC that are too large or the model has too few conformations. 13C13 NMR relaxation data on this polysaccharide lead us to suspect the latter defect to be the source of the discrepancy [29]. NOE data are not very sensitive to details of the model since simulated NOE from the model of Xu and Bush [19] agree fairly well with the experimental data. Better validation of the 3JCH and 3JCC coupling constant correlation curves is needed including more data on relatively rigid oligosaccharides such as blood group epitopes for which one can be confident about the model conformation.
ACKNOWLEDGMENT
We thank Dr. M. F. Summers for providing the observation time on the G.E. Omega 600 spectrometer.
References itemize
Table 1.
nJCC measured for uniformly 13C13 enriched glucose (in Hz.)
| Carbon | a-glc | b-glc | ||
| meas. | lita | meas | lit | |
|
C1-C3 | 0 | 0 | 4.5 | 4.6 |
| C1-C4 | 0 | 0 | 0 | 0 |
| C1-C5 | 2.1 | 1.8 | olb C3 | 0 |
| C1-C6 | 3.4 | 3.3 | 4.0 | 4.1 |
| C3-C6 | ol | 3.9 | 4.5 | 4.4 |
a.) Literature values from King-Morris and Serianni [21] and Wu et al., [22]
b.) ol indicates that overlapping cross peaks prevent accurate measurement.
Table 2.
nJCC (Hz) for the Polysacchairde of S. mitis J22
| Carbon | a | b | g | c | d | e | f | ||||||
| meas. | lita | meas | lit | meas | lit | meas | lit | meas | meas | lit | meas | lit
| |
| C1-C3 | 0c | 0 | 5.05 | 4.0 | ol C2 | 0 | 4.25 | 4.5 | 3.42 | 4.9 | 4.6 | 0 | 0 |
|
C1-C4 | 0 | 0 | 0 | 0 | ol Cx | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| C1-C5 | 1.64 | 1.9 | 0 | 0 | 1.53 | 2.0 | 0 | 0 | 1.79 | 0 | 0 | 1.98 | 1.9 |
|
C1-C6 | 3.48 | 3.6 | limb | lim | 3.68 | 4.1 | - | 3.89 | 4.4 | 3.57 | 3.6 | ||
|
C1-Cx | 1.77 | 1.80 | ol | 1.40 | 1.70 | 2.04 | - | ||||||
| C1-Cx-1 | 2.27 | 0 | ol C4 | 2.19 | 2.48 | lim | - | ||||||
| C1-Cx+1 | 2.49 | 1.94 | 0 | 0? | - | ol | - | ||||||
| C2-Cx | 1.73 | ol C2 | ol aC5 | phase | 2.61 | ol | - |
a.) Literature values from King-Morris and Serianni (1987)
b.) lim. indicates that the 13C13 chemical shift difference is too large for simultaneous excitation with our instrument.
c.) 0 means that nJCC is less than 1 Hz in our data and less than 0.8 Hz in data of King-Morris and Serianni (1987)
Figure 2. 2-d quantitative coherence transfer correlation spectrum of the lectin-receptor polysaccharide from S. mitis J22. Columns at the 1H1 resonances of residue e b-gal which is linked (1\rightarrowfill3)-to residue f a-galNAc. Strong cross peaks correspond to large coupling constants to C3 and C6 of that residue and to C3 of f. The resonance of f C2 (d=49.5 ppm) is too far from e C1 for simultaneous 90°excitation and the weak peaks to f C4 and e C4 overlap. Column at the 1H1 resonance of residue c shows cross peaks corresponding to coupling of c C1 (b-glucose) with large coupling constants to C3 and C6 and smaller coupling constants to d C6 (linkage position) and to d C5 (Cx-1), (See ref [25] for 1H1 and 13C13 chemical shift assignments.)
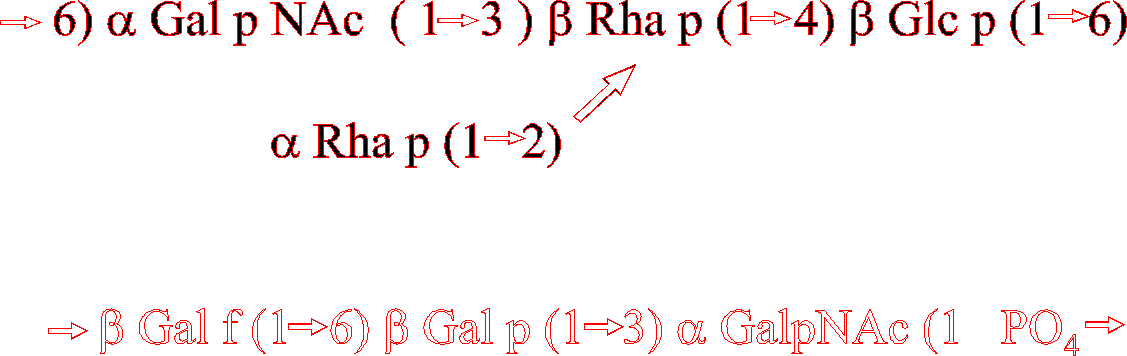{kind=link}
{kind=link}